
Фолікулярні Т-хелпери
Фолікулярні Т-хелпери (також відомі як фолікулярні Т-хелпери В-клітин та скорочено TFH) є антиген-активованими CD4+ Т-клітинами, які знаходяться всередині В-клітинних фолікулів вторинних лімфоїдних органів, таких як лімфатичні вузли, селезінка та Пеєрові бляшки, і ідентифікуються за їх конститутивною експресією рецептора хомінгу В-клітинного фолікула CXCR5.[1] Після клітинної взаємодії та перехресної сигналізації зі спорідненими фолікулярними В-клітинами, TFH запускають утворення та підтримку зародкових центрів за допомогою експресії ліганду CD40 (CD40L) та секреції IL-21 [2] та IL -4 . [3] ТFH також мігрують із Т-клітинних зон до цих осередків зародкових центрів, які переважно складаються з В-клітин, що швидко діляться і мутують. Усередині зародкових центрів TFH відіграють важливу роль у посередництві відбору та виживання В-клітин, які потім диференціюються або в довгоживучі плазматичні клітини, здатні продукувати високоафінні антитіла проти чужорідного антигену, або в В-клітини пам'яті, здатні до швидкої реактивації при повторній зустрічі з тим же антигеном.[4] Вважається також, що TFH сприяють негативному відбору потенційно аутоімунних мутованих В-клітин у зародковому центрі. Однак механізми, за допомогою яких ТFH опосередковують толерантність зародкового центру, ще не повністю зрозумілі.
Цілком можливо, що TFH можуть виникати як розгалуження в шляхах диференціювання Th1 і Th2, але їх точне відношення до інших ефекторних субпопуляцій CD4+ Т-клітин все ще залишається невизначеним. Однак дослідження показали, що TFH мають різні профілі експресії генів, що підтверджує теорію, що TFH є підгрупою CD4+ T клітин, відмінних від Th1, Th2, Th-17 або Tregs . [5] [6]
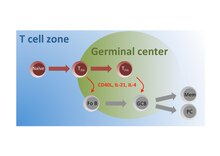
Підгрупа наївних Т-клітин у Т-клітинній зоні активується антигеном і мігрує до фолікулів, де вони диференціюються в ТFH, які взаємодіють з фолікулярними B-клітинами (Fo B) та "інструктують" їх на перемикання класу імуноглобулінів, соматичній гіпермутації та швидкий клітинний розвитоку, поділ для "засіювання" зародкових центрів. У цих зародкових центрах TFH продовжують надавати допомогу В-клітинам, щоб полегшити їх перетворення на плазматичні клітини що продукують високоафінні антитіла і на В-клітини довготривалої пам’яті.
Доведено, що костимулятор Т-клітин (CD278 або ICOS) є особливо критичним сигналом для ТFH, так як у мишей з дефіцитом в ICOS TFH не утворюються.[7] Крім того, було показано, що ICOS індукує секрецію цитокіну IL-21 активованими CD4 + Т-клітинами і що IL-21 відіграє вирішальну роль у розвитку TFH- клітин і зародкових центрів.[8] [9] Також Bcl-6 є фактором транскрипції, ідентифікованим у TFH, але він може мати інші ролі за межами цієї підгрупи, оскільки він також бере участь у розвитку CD8+ Т-клітин пам’яті.[10]
У зародкових центрах антиген-активані ТFH, швидко посилюють експресію CD40L, який зв’язує і стимулює поверхневий рецептор В-клітин CD40.[11] TFH паракринна активація В-клітин CD40 призводить до виживання та диференціювання В-клітин, включаючи індукцію AID ( індукованої активацією (цитидин) деамінази).[12] Експресія AID (кодується геном AICDA ) змушує В-клітинні антитіла перемикатися з IgM/IgD на інші ізотипи антитіл і викликає соматичну гіпермутацію під час клональної проліферації. Змінені антитіла набувають кращих ефекторних функцій, а гіпермутовані антитіла демонструють більшу спорідненість до антигену.
TFH, утворені на ранніх стадіях реакції зародкового центру, офіційно називаються пре-TFH. Вони унікально зустрічаються переважно на кордоні Т-клітинної зони, яка зливається з В-клітинними фолікулами і зародковими центрами. Пре-TFH функціонально дуже подібні до інших TFH у сприянні реакцій В-клітин зародкового центру, однак, крім того, вони також здатні стимулювати перетворення фолікулярних В-клітин поблизу та поза зародковими центрами плазматичні клітини, які реагують швидким, але не тривалим виробленням антитіл (що відоме як екстрафолікулярна відповідь).
Ті специфічні ТFH які проживають в межах зрілого зародкового центру іноді називають GC ТFH (ТFH зародкових центрів) щоб відрізнити їх від клітин пре-ТFH.[13] [14] У цій популяції також виділяють субпопуляцію GC Tfh - клітини, які експресують ген Foxp3, що кодує фактор транскрипції. Ця невелика окрема субпопуляція клітин, яка TFR (для фолікулярних регуляторних Т-клітин), важлива для допомоги в контролі та обмеженні величини нормальних реакцій зародкового центру таким чином, щоб не вироблялися аномально мутовані або самореактивні антитіла (аутоантитіла).[15] Таким чином, TFR є унікальним гальмівним впливом під час реакції зародкового центру.
У той час як ТFH знаходяться в основному у вторинних лімфоїдних органах, невелика їх частка циркулює в крові і називається «периферичними» (pTFH). Ці клітини можна ідентифікувати за експресією IL-21 після стимуляції. [16]
ТFH вважаються незамінною підгрупою Т-клітин у генерації та підтримці реакцій зародкового центру. Таким чином, за відсутності ТFH, подібно до активації В-клітин незалежними від Т-клітин антигенами, утворюється швидкий сплеск продукції плазматичних клітин, які продукують низькоафінні антитіла, але це не призводить до індукції зародкового центру і не дозволяє відбутися дозріванню афінності антитіл або диференціації ефективних В-клітин пам’яті, які необхідні для вироблення стійкості організму проти повторних інфекцій. Зокрема, В-клітини пам’яті, що залежать від зародкового центру, є драйверами повторного вироблення антитіл під час вторинної імунної відповіді. Таким чином, правильна активація та розвиток TFH є центральним елементом ефективності імунізації та розробки вакцини для індукції довгострокового імунітету. У популяційному дослідженні в Бангладеш за участі пацієнтів, інфікованих холерним вібріоном, і здорових людей-добровольців, яким вводили наявну холерну вакцину[17] реакція пам’яті TFH, специфічна проти холерного антигену, корелювала з подальшим виробленням антитіл В-клітинами.
З нормальним старінням відбувається поступове зниження активності імунної системи організму. Це явище, яке називається імуностарінням, значною мірою пов’язане зі зниженням функції Т-клітин, включаючи здатність TFH належним чином підтримувати реакції зародкового центру.[18] Частково це може бути пов’язано з нижчим рівнем CD40L на клітинній поверхні ТFH у літніх.[19]
Неконтрольовані або надмірно активні імунні відповіді TFH можуть призвести до появи неконтрольованих зародкових центрів, що складаються з аберантно мутованих В-клітин, які можуть викликати опосередковані антитілами аутоімунні захворювання. Підвищені рівні TFH-подібних клітин можна виявити в крові частини пацієнтів із системним червоним вовчаком і синдромом Шегрена.[20] Однак наукові докази, які свідчать про те, що ТFH точно можуть викликати аутоімунні реакції у людей, залишаються неповними.
- ↑ Fazilleau; Mark, L; McHeyzer-Williams, LJ; McHeyzer-Williams, MG та ін. (March 2009). Follicular helper T cells: lineage and location. Immunity. 30 (3): 324—35. doi:10.1016/j.immuni.2009.03.003. PMC 2731675. PMID 19303387.
- ↑ Seo; Youn, J; Kim, PH та ін. (May 2009). IL-21 ensures TGF-beta 1-induced IgA isotype expression in mouse Peyer's patches. J Leukoc Biol. 85 (5): 744—50. doi:10.1189/jlb.0708450. PMID 19168593.
- ↑ Zaretsky; Taylor, JJ; King, IL; Marshall, FA; Mohrs, M; Pearce, EJ та ін. (May 2009). T follicular helper cells differentiate from Th2 cells in response to helminth antigens. J Exp Med. 206 (5): 991—9. doi:10.1084/jem.20090303. PMC 2715032. PMID 19380637.
- ↑ Kurosaki; Kometani, K; Ise, W (March 2015). Memory B cells. Nat Rev Immunol. 15 (3): 149—59. doi:10.1038/nri3802. PMID 25677494.
- ↑ Chtanova; Tangye, SG; Newton, R; Frank, N; Hodge, MR; Rolph, MS; MacKay, CR та ін. (July 2004). T follicular helper cells express a distinctive transcriptional profile, reflecting their role as non-Th1/Th2 effector cells that provide help for B cells. J Immunol. 173 (1): 68—78. doi:10.4049/jimmunol.173.1.68. PMID 15210760.
- ↑ Nurieva RI, Chung Y, Hwang D, Yang XO, Kang HS, Ma L, Wang YH, Watowich SS, Jetten AM, Tian Q, Dong C (2008). Generation of T follicular helper cells is mediated by interleukin-21 but independent of T helper 1, 2, or 17 cell lineages. Immunity. 29 (1): 138—49. doi:10.1016/j.immuni.2008.05.009. PMC 2556461. PMID 18599325.
{{cite journal}}: Недійсний|displayauthors=8(довідка) - ↑ Akiba; Takeda, K; Kojima, Y; Usui, Y; Harada, N; Yamazaki, T; Ma, J; Tezuka, K та ін. (Aug 2005). The role of ICOS in the CXCR5+ follicular B helper T cell maintenance in vivo. J Immunol. 175 (4): 2340—8. doi:10.4049/jimmunol.175.4.2340. PMID 16081804.
- ↑ Bauquet; Jin, H; Paterson, AM; Mitsdoerffer, M; Ho, IC; Sharpe, AH; Kuchroo, VK (Feb 2009). The costimulatory molecule ICOS regulates the expression of c-Maf and IL-21 in the development of follicular T helper cells and TH-17 cells. Nat Immunol. 10 (2): 167—75. doi:10.1038/ni.1690. PMC 2742982. PMID 19098919.
- ↑ Vogelzang; McGuire, HM; Yu, D; Sprent, J; MacKay, CR; King, C (July 2008). A fundamental role for interleukin-21 in the generation of T follicular helper cells. Immunity. 29 (1): 127—37. doi:10.1016/j.immuni.2008.06.001. PMID 18602282.
- ↑ Ichii; Sakamoto, A; Hatano, M; Okada, S; Toyama, H; Taki, S; Arima, M; Kuroda, Y; Tokuhisa, T та ін. (June 2002). Role for Bcl-6 in the generation and maintenance of memory CD8+ T cells. Nat Immunol. 3 (6): 558—63. doi:10.1038/ni802. PMID 12021781.
- ↑ Kim, JR; Lim, HW; Kang, SG; Hillsamer, P; Kim, CH (4 лютого 2005). Human CD57+ germinal center-T cells are the major helpers for GC-B cells and induce class switch recombination. BMC Immunology. 6: 3. doi:10.1186/1471-2172-6-3. PMC 548684. PMID 15694005.
{{cite journal}}: Обслуговування CS1: Сторінки із непозначеним DOI з безкоштовним доступом (посилання) - ↑ McHeyzer-Williams, M; Okitsu, S; Wang, N; McHeyzer-Williams, L (9 грудня 2011). Molecular programming of B cell memory. Nature Reviews. Immunology. 12 (1): 24—34. doi:10.1038/nri3128. PMC 3947622. PMID 22158414.
- ↑ Yusuf I, Kageyama R, Monticelli L, Johnston RJ, Ditoro D, Hansen K, Barnett B, Crotty S (July 2010). Germinal center T follicular helper cell IL-4 production is dependent on signaling lymphocytic activation molecule receptor (CD150). J. Immunol. 185 (1): 190—202. doi:10.4049/jimmunol.0903505. PMC 2913439. PMID 20525889.
- ↑ Ramiscal RR, Vinuesa CG (March 2013). T-cell subsets in the germinal center. Immunol. Rev. 252 (1): 146—55. doi:10.1111/imr.12031. PMID 23405902.
- ↑ Wollenberg I, Agua-Doce A, Hernández A, Almeida C, Oliveira VG, Faro J, Graca L (November 2011). Regulation of the germinal center reaction by Foxp3+ follicular regulatory T cells. J. Immunol. 187 (9): 4553—60. doi:10.4049/jimmunol.1101328. PMID 21984700.
- ↑ Schultz, Bruce T.; Teigler, Jeffrey E.; Pissani, Franco; Oster, Alexander F.; Kranias, Gregory; Alter, Galit; Marovich, Mary; Eller, Michael A.; Dittmer, Ulf (19 січня 2016). Circulating HIV-Specific Interleukin-21(+)CD4(+) T Cells Represent Peripheral Tfh Cells with Antigen-Dependent Helper Functions. Immunity. 44 (1): 167—178. doi:10.1016/j.immuni.2015.12.011. ISSN 1097-4180. PMID 26795249.
- ↑ Arifuzzaman M, Rashu R, Leung DT, Hosen MI, Bhuiyan TR, Bhuiyan MS, Rahman MA, Khanam F, Saha A, Charles RC, LaRocque RC, Weil AA, Clements JD, Holmes RK, Calderwood SB, Harris JB, Ryan ET, Qadri F (August 2012). Antigen-specific memory T cell responses after vaccination with an oral killed cholera vaccine in Bangali children and comparison to responses in patients with naturally acquired cholera. Clinical and Vaccine Immunology. 19 (8): 1304—11. doi:10.1128/CVI.00196-12. PMC 3416086. PMID 22739692.
- ↑ Lefebvre JS, Maue AC, Eaton SM, Lanthier PA, Tighe M, Haynes L (2012). The aged microenvironment contributes to the age-related functional defects of CD4 T cells in mice. Aging Cell. 11 (5): 732—40. doi:10.1111/j.1474-9726.2012.00836.x. PMC 3444657. PMID 22607653.
- ↑ Perkey E, Miller RA, Garcia GG (2012). Ex vivo enzymatic treatment of aged CD4 T cells restores cognate T cell helper function and enhances antibody production in mice. J Immunol. 189 (12): 5582—9. doi:10.4049/jimmunol.1200487. PMC 3518580. PMID 23136198.
- ↑ Craft JE. (2012). Follicular helper T cells in immunity and systemic autoimmunity. Nat Rev Rheumatol. 8 (6): 337—47. doi:10.1038/nrrheum.2012.58. PMC 3604997. PMID 22549246.